 2023, Vol. 41
2023, Vol. 41Institute of Oceanology, Chinese Academy of Sciences
Article Information
- DENG Dailan, MENG Han, MA You, GUO Yongqi, WANG Zixuan, HE Huan, WAQAS Khan, LIU Jin'e, ZHANG Limin
- Effect of extracellular polymeric substances on Dolichospermum aggregation during temperature rise
- Journal of Oceanology and Limnology, 41(6): 2208-2218
- http://dx.doi.org/10.1007/s00343-023-2249-8
Article History
- Received Jun. 15, 2022
- accepted in principle Aug. 17, 2022
- accepted for publication Nov. 25, 2022




2 Green Economy Development Institute, Nanjing University of Finance and Economics, Nanjing 210023, China;
3 Jiangsu Engineering Lab of Water and Soil Eco-remediation, Nanjing 210023, China
With the increasing eutrophication in freshwater lakes, cyanobacteria blooms have become a global environmental problem, which occurred commonly in eutrophic lakes, ponds, rivers besides threatened the sustainability of freshwater ecosystems (Paerl and Paul, 2012). Temperature is considered as an important factor that directly controls the distribution and productivity of algae (Holsinger, 1955; Sakharova et al., 2020). Since the 20th century global cyanobacterial blooms have broken out widely (Carey et al., 2012; Maheaux et al., 2016).
In recent years, filamentous cyanobacteria blooms have occurred recurrently around the world, for example, Poyang Lake in China (Qian et al., 2019), the Baltic Sea in Northern Europe (Olofsson et al., 2019), Lake Balaton in Central Europe (Kovács et al., 2016), and Lake Sonachi and Lake Simbi in Africa (Ndlela et al., 2016). There are many environmental factors affecting the growth of filamentous cyanobacteria, among which, temperature is one of the main environmental factors. The fact that cyanobacteria preferred warm temperatures, many studies have been conducted to investigate cyanobacteria expansion might be changed by global warming (Paerl et al., 2011; Carey et al., 2012; Maheaux et al., 2016). Consequently, it remains to note that the duration, distribution, and intensity of cyanobacterial blooms are probably increased with increasing temperatures (Paerl and Huisman, 2009).
Dolichospermum is a photoautotrophic filamentous cyanobacterium capable of fixing atmospheric nitrogen (Zhang et al., 2021). In appropriate conditions, hundreds of Dolichospermum cells directly connect (Arévalo et al., 2021) and form filaments. These filaments could rapidly response to adapt to environmental changes (Weiss et al., 2019). For example, Dolichospermum cells form spore-like resting cells in cold winters (Garg and Maldener, 2021). They are dormant cells which different from vegetative cells, helping bacteria to survive under severely cold environments (Sukenik et al., 2019). When the temperature is appropriate, the dormant organism recovers, reproduces and form aggregates (Garg and Maldener, 2021; Srivastava et al., 2021). It is reported cyanobacteria quickly resume their metabolism and photosynthetic rates when lake ice melted (Olson et al., 1998; Paerl et al., 1998). Furthermore, Reddy et al. (2019) discovered that Dolichospermum is sensitive to high temperature. Elevated temperature could inhibit growth and photosynthesis of Dolichospermum, produce reactive oxygen species, and damage RNA and protein (Wen et al., 2005). However, a large number of studies on the growth of Dolichospermum focused on the culture at a single temperature, and few studies conducted at a continuous temperature.
In addition, the direct cause of algal blooms is supposed to be the accumulation of algal cells, which promotes vertical migration and rises to the lake surface to form algal collections (Chen et al., 2019). The microbial collection is largely influenced by surface properties, including zeta potential and hydrophobicity/hydrophilicity (Sheng et al., 2010). As the extracellular polymeric substances (EPSs) directly covered with the surface of algae cells, it is a kind of polymer organic matter produced through adsorption, excretion, secretion, and cell lysis. EPSs are composed of polysaccharides, proteins, lipids, and humus (Xu et al., 2010, 2013a; Liu et al., 2017). The polysaccharides were initially considered the main components of cyanobacteria EPSs (Forni et al., 1997). More recently, protein-corresponding substances have been found in EPSs (Xu et al., 2013a; Liu et al., 2017). The content and composition of EPSs were supposed to enhance cyanobacteria proliferation through cell adhesion and other processes (Yang et al., 2008), and can also alter the surface properties of algae cells to promote algae cells aggregation (Liu and Fang, 2002; Vogelaar et al., 2005; Yu et al., 2009).
Furthermore, EPSs were divided into three fractions, including soluble EPS (S-EPS), loosely bound EPS (LB-EPS), and tightly bound EPS (TB-EPS) (Xu et al., 2013a). Xu et al. (2013b) found that most organic matter was dispersed in TB-EPS, and only a small amount was dispersed in S-EPS and LB-EPS. Among them, TB-EPS played role in the process of cyanobacteria combination and stimulates the formation of algal cell collections, while LB-EPS contributes to the development of the combinations towards the direction of bloom (Xu et al., 2014).
Despite numerous researches followed on EPSs components and found soluble and binding EPSs secreted by cyanobacteria at certain growth stages have effects on algae growth (Yang et al., 2008; Qu et al., 2012b). But most of them focused on Microcystis (Xu et al., 2013a, b, 2014), and less attention has been paid to filamentous cyanobacteria, for example, Dolichospermum. Therefore, the dynamic of EPSs separation process, response to the growth and aggregation of filamentous cyanobacteria cells remain unclear.
Dolichospermum is a harmful alga that produces anatoxins, which are more harmful than microcystin (Swe et al., 2021). Moreover, Dolichospermum, the heterotrophic nitrogen-fixing microorganism, adverse environments (Zhang et al., 2021). Thus, clarifying the growth and aggregation mechanism of Anabaena will contribute to preventing algal blooms, and protecting the health of the environment.
In this study, Dolichospermum was studied in laboratory. Dolichospermum was cultivated under temperature rise treatments, and its growth dynamics and aggregation characteristics were determined. The EPSs released and aggregation characteristics, including contact angles and zeta potential, were systematically determined. The surface thermodynamic analysis was applied to interpret the results and the roles of EPSs, and explored its subfractions in Dolichospermum aggregation. The results can help explain the aggregation change of Dolichospermum in aquatic environment under temperature change, and provide a basis for the management of filamentous cyanobacteria.
2 MATERIAL AND METHOD 2.1 Cultivation of DolichospermumThe cyanobacterial strain Dolichospermum (PCC 7120), was provided by the Institute of Hydrobiology, Chinese Academy of Sciences, and it was cultured in conical flasks with batch mode. Before cultivation, all experimental utensils and mediums were put in autoclaved at 121 ℃ for 30 min. Dolichospermum in exponentially growing stage was injected into a conical flask containing 1 200-mL BG-11 medium, and the initial absorbance was adjusted to 0.1, then evenly divided into three conical flasks and cultured at a light꞉dark cycle of 12 h꞉12 h at a light intensity of 30 μmol photons/(m2·s). Dolichospermum growth was determined through taking absorbance of Dolichospermum cultures at 680 nm using a spectrophotometer (Tecan SPARK, Austria). To simulate a temperature change gradient, Dolichospermum were first acclimated at 7 ℃ for a week to adapt the low-temperature environment, and then the culture temperature increased by 2 ℃ every 3 days from 7 ℃ to 37 ℃. The same volumes of algal fluid samples were collected to analyze on the last day of each temperature setting. All the flasks were shaken by hand three times a day at regular intervals during the experimental period. Each treatment was conducted in triplicate.
2.2 EPSs extraction and determination of fluorescence EEMEPSs consist of S-EPS, LB-EPS, and TB-EPS (Li and Yang, 2007; Sheng et al., 2010; Xu et al., 2010). In this study, EPSs were extracted according to Xu et al. (2013a). All cuvettes were rinsed before being analyzed. The three-dimensional excitation-emission matrix fluorescence spectrum (3D-EEM) was measured using a fluorescence spectrometer (LF-1301009, Thermo Fisher), and the measurement was carried out in accordance with Xu et al. (2013b). The difference is that the emission (Em) and excitation (Ex) wavelengths range from 250 nm to 450 nm at 5-nm increments and from 200 nm to 550 nm at 10-nm increments, respectively. Further, Rayleigh and Raman scattering were filtered using interpolation adopted from Bahram et al. (2006).
2.3 Measurement of zeta potential and contact angleThe zeta potential represents the electrostatic force on the cell surface, with which the stability of dispersions can be evaluated (Tan et al., 2020). In this study, the zeta potential was measured at 25 ℃ using a Zeta sizer (Nano ZS90, Malvern Instruments Ltd., UK) according to Hadjoudja et al. (2010). Before the test, the collected 50-μL sample was diluted 100 times with PBS buffer solution. All measurements were conducted in triplicate.
The contact angle (θ) was measured using a contact angle analyzer (JC2000D, Powereach, Shanghai, China) according to the standard sessile drop method (Funke, 1995). Pure water, glycerol, and diiodomethane were dropped onto the membrane containing Dolichospermum, separately. Each sample was tested at least seven times to obtain the arithmetic mean.
2.4 Interfacial thermodynamic characteristics of DolichospermumThe changes in the interfacial thermodynamics of Dolichospermum surface in different temperatures were estimated by contact angle measurement. The interfacial free energy (ΔGslIFE), where sl means sample and liquid, between the algae and water is the sum of van der Waals interaction free energy (ΔGslLW) and Lewis acid-base interaction free energy (ΔGslAB) (Florence, 2007). The value of ΔGslIFE can characterize the interaction between algae and water (Hou et al., 2015). When ΔGslIFE > 0, the binding force between algae and water is stronger than that between water molecules, and algae cells are more inclined to combine with water molecules than with each other, so the algae exhibit hydrophilicity. The detailed calculation process is described in Supplementary equations 1–11 and the relevant parameters involved were shown in Supplementary Table S1.
2.5 Dolichospermum growth detection and determination of aggregation abilityCell density was measured at absorbance of 680 nm (OD680). Furthermore, the maximum photoelectron production (Fv/Fm) and actual photoelectron production (Fv′/Fm′) were measured using a pulse amplitude modulated fluorometer (AquaPen, PSI, Czech Republic).
The aggregation ability of the algal cells is usually evaluated by the aggregation rate (Tan et al., 2018). The aggregation rate was determined according to Xu et al. (2014). At 680 nm, the optical density of Dolichospermum sample was determined (A0). After 6 h of standing in a clean test tube, the algal liquid from 2 cm of the upper layer was carefully removed and the optical density was measured again at 680 nm (At). The aggregation ratio of Dolichospermum samples was calculated by using the following equation:
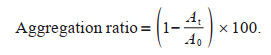
All chemicals used in this work were of analytical grade. Data are presented as the means± standard deviations. Significant differences were analyzed by one-way analysis of variance (ANOVA). All statistical analyses were performed by SPSS 22.0. Pearson correlation and principal component analysis (PCA) were performed to describe the relationships among the EPS content, aggregation ability, and zeta potential.
The polysaccharide content was determined using the phenol sulfuric acid with glucose as a standard (Dubois et al., 1956). Protein content was measured according to Bistgani et al. (2017) using bovine serum albumin as standard.
3 RESULT 3.1 Changes of growth and aggregation ability of DolichospermumAt 7–11 ℃, photosynthetic activity entered resting status. Fv/Fm did not increase significantly, and the biomass (OD680 valua) remained unchanged at 7–13 ℃. With the recovery of the photosynthetic activity at 11 ℃ (Fig. 1a), Dolichospermum biomass began to increase significantly at 13 ℃ (Fig. 1b). Thus, the recruitment temperature of Dolichospermum was determined as 11 ℃ in this study. In addition, the lower limit of temperature tolerance was 11 ℃ for Dolichospermum growth.

|
| Fig.1 Effects of temperature on the growth of Dolichospermum and aggregation variations a. variation in photosynthetic activity of Dolichospermum with temperature; b. variation in algal biomass with temperature; c. variation in the aggregation rate of Anabaena with temperature. Fv/Fm: the maximum photoelectron production; Fv′/Fm′: the actual photoelectron production; OD680: the absorbance in 680 nm. |
Then, when the cultivation temperature was improved to 21 ℃, the maximum photoelectron production (Fv/Fm), actual photoelectron production (Fv′/Fm′) and the growth rate reached the maximum. Therefore, the optimum temperature for Dolichospermum growth was considered 21 ℃ in this experiment.
When the cultivation temperature kept increasing from 21 ℃ to 37 ℃, the photosynthetic activity and the growth rate decreased. Finally, the biomass stopped increasing at 35 ℃, which was considered the upper limit of temperature tolerance for Dolichospermum growth.
At 7–11 ℃, the aggregation rate increased rapidly, while the aggregation rate decreased significantly at 11–37 ℃ (Fig. 1c). It was observed that the aggregating of Dolichospermum occurred at the stage of rapid growth when the photosynthetic activity and the growth rate increased rapidly.
It may be the protein that promoted Dolichospermum aggregation in EPSs components because the significant increase of the protein concentration in the EPSs was observed at 11–21 ℃ (Fig. 2b). It was proved that most polysaccharides exhibited hydrophilicity and proteins exhibited hydrophobicity (Liu et al., 2010). When the protein proportion increased, Dolichospermum tended to be hydrophobic and easier to gather. As the temperature continued to increase, the protein proportion decreased. Consequently, the algal cells tended to be hydrophilic and difficult to aggregate.

|
| Fig.2 Variations of polysaccharides and proteins in EPSs matrix with increasing temperature a. concentration of polysaccharide content in EPSs components; b. concentration of protein content in EPSs components. R2: pearson correlation coefficient; **: P < 0.01. |
Extracellular polymers enhance the ability of algae to resist external stress (Dang et al., 2018; Wang et al., 2018). The extracellular polysaccharide concentration increased with increasing temperature, and the extracellular protein concentration was increased and reached the maximum value at 23 ℃ (Fig. 2). The change rates of polysaccharides and proteins released by Dolichospermum in high temperatures (27–37 ℃) were higher than that in low temperatures (7–15 ℃). In addition, the concentrations of protein in EPSs were always lower than that of polysaccharides with the temperature rising during the growth period, indicating that the Dolichospermum EPSs consisted of a major fraction of polysaccharides. This is consistent with the results reported in a lab-cultivated study of M. aeruginosa (Qu et al., 2012a; Xu et al., 2013b).
The polysaccharide concentration of the three EPSs fractions increased with the temperature rise, while the protein concentration increased before 23 ℃ and then decreased (Fig. 2). This might have correlation to Dolichospermum tolerance to temperature that, high temperatures damaged protein structures (Wen et al., 2005). The polysaccharide concentration in S-EPS increased slowly at low temperatures (7–15 ℃), and increased when the temperature was higher than 19 ℃. Then, it reached the maximum at 37 ℃. However, the polysaccharide concentrations in LB-EPS and TB-EPS were lower than that in S-EPS and hardly influenced. The protein concentration of the three EPSs fractions increased with increasing temperature until the temperature was higher than 25 ℃, indicating that the synthesis of some proteins in Dolichospermum was inhibited at the high temperature (> 25 ℃). Furthermore, the concentrations of protein in S-EPS and LB-EPS were higher than that of TB-EPS. Some researchers reported that in Microcystis EPSs, the TB-EPS and S-EPS mainly consisted of polysaccharides, and LB-EPS were mainly composed of proteins (Xu et al., 2013b). In this study, polysaccharides are dispersed in S-EPS mainly, and proteins are dispersed in mainly S-EPS and LB-EPS, indicating that filamentous cyanobacteria had a different EPSs composition from unicellular cyanobacteria.
Principal component analysis (PCA) showed that the two axes explained 76.4% of the total variance of the aggregation of Dolichospermum (Fig. 3). It indicated that the aggregation rates were positively correlated to the proteins in EPSs (especially S-EPS and LB-EPS proteins) and the zeta potential. However, the extracellular polysaccharides were negatively related to the aggregation.

|
| Fig.3 Principal component analysis of variables related to Dolichospermum aggregation process OD680: the absorbance in 680 nm; Fv/Fm: the maximum photoelectron production; Chl a: chlorophyll a; S-EPS: soluble extracellular polymeric substances; LB-EPS: loosely bound extracellular polymeric substances; TB-EPS: tightly bound extracellular polymeric substances. The red circle: 95% confidence ellipse. |
EEM is widely used in the characterization of organic matters. Because it could obtain intensity information of wavelength with the advantages of high sensitivity (He et al., 2017). The park A and park B were the humus like substances located at Ex/Em of 360 nm/450 nm and Ex/Em of 320 nm/450 nm, respectively (details about the data of peaks in Supplementary Fig.S1). The peak C was the protein like substances located at Ex/Em of 290 nm/350 nm. The fluorescence is related to tryptophan protein-like substances (Baker and Inverarity, 2004). Both humus-like substances (peaks A and B) and protein-like substances (peaks C) were observed in the S-EPS and LB-EPS, and only protein-like substances (peaks C) were observed in TB-EPS (Fig. 4). It means that the TB-EPS of Dolichospermum was composed of protein-like substances. Protein is an important component of cyanobacteria cells and can be released during cell metabolism. Henderson et al. (2008) reported the presence of proteins in algae-producing organisms extracted from cyanobacteria, green algae, and diatoms. Humus-like substances may originate from the decomposition of dead cells and macromolecular organics such as proteins and polysaccharides (Qu et al., 2012a). Similar fluorescence peak positions were detected in the three EPSs fractions at different temperatures, showing that each kind of EPSs was not influenced by temperature. Nevertheless, the fluorescence intensity of EPSs fractions was different with increasing temperature, implying that the concentration of specific compounds in each kind of EPSs was affected by temperature.

|
| Fig.4 The EEM contours of different EPSs fractions at 9, 17, and 25 ℃ for Dolichospermum a, b, and c. S-EPS; d, e, and f. LB-EPS; g, h, and i. TB-EPS; peaks A, B, and C were three peaks located at Ex/Em of 360 nm/450 nm, 320 nm/450 nm and 290 nm/350 nm, respectively. |
To reveal the influence of physicochemical properties of algae on the aggregation, the contact angles and zeta potential were analyzed (Table 1). The contact angle between a sample and water could characterize the hydrophobicity of Dolichospermum. Results show that Dolichospermum was characterized as being hydrophilic (θH2O≤90°).

|
The zeta potential indicated the electrostatic repulsion on the surface of the algae, and the higher zeta potential value represented the less aggregation capacity. The absolute value of zeta potential increased with increasing temperature, which was consistent with the trend of the total concentration of EPSs, details described in Supplementary Fig.S2. This indicated that the EPSs secretion of Dolichospermum increased, the negative charge on the surface of algal cells, and enhanced the repulsion between the algal cells.
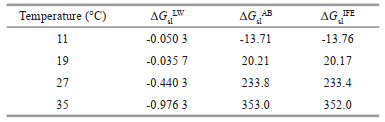
3.3.2 Surface thermodynamic analysis of DolichospermumSurface thermodynamic analysis was applied to examine the aggregation ability of Dolichospermum (Table 2). The value of ΔGslIFE increased from -13.76 to 352.0 mJ/m2 during the cultivation, indicating the decrease of surface hydrophobicity. Since the Lifshitz-van-der-Waals interaction force is always attractive, the values of ΔGslLW were always negative, which was following the results reported by Florence (2007). However, the value of ΔGslAB was greater than that of ΔGslLW at each temperature, implying that ΔGslAB was the major contributor to ΔGslIFE. The Lewis acid-base interactions played a key role in the aggregation.
The effective Hamaker constant (ASLS) value represented the intermolecular attraction (Funke, 1995; Liu et al., 2007). The electrostatic force (WEI) was mainly influenced by surface charge. In this study, with temperature increase, the ASLS value increased, and the WEI value gradually increased too (Fig. 5). Results show that with temperature increase, the surface charge quantity of Dolichospermum increased, the electrostatic repulsion on the surface of algae cells enhanced, the inter-particle attraction decreased, so the algae cells tended to disperse instead of aggregating, which is consistent with the results of EPSs (Fig. 2) components and zeta potential.

|
| Fig.5 Changes of the effective Hamaker constant (a) and the electrostatic force (b) |
Polysaccharides and proteins were the main components of EPSs, accounting for 70%–80% of the total EPSs (Richert et al., 2005), and most polysaccharides had hydrophilic properties, while proteins were more hydrophobicity (Liu et al., 2010). Previous studies (Liu et al., 2014; Ji et al., 2021) and this experiment showed that bound EPSs (LB-EPS and TB-EPS) had more hydrophobic components and fluorescent components than S-EPS. Therefore, it can be concluded that extracellular polysaccharides inhibited the aggregation, while protein promoted the aggregation of Dolichospermum.
Furthermore, surface charge was the key factor affecting aggregation (Xing et al., 2022). If the surface negative charge between the cyanobacteria particles was large enough, the repulsion force between the particles was strong and difficult to aggregation. Tan et al. (2018) found that the increase of the ratio of protein to polysaccharide would reduce the net charge on the surface of algae cells, and then reduced the electrostatic repulsion. In addition, proteins are charge neutralizers because some proteins with positively charged groups could neutralize anionic functional groups (Liao et al., 2001). Moreover, the ratio of protein to polysaccharide was positively correlated with hydrophobicity, which is consistent with the conclusion obtained from the surface thermodynamic analysis in this study.
Above all, it could be explained that the protein of EPSs promoted the aggregation of Dolichospermum because it reduced the net charge on the surface of algae cells, and then reduced the electrostatic repulsion. In addition, when the polysaccharide of EPSs increased, the aggregation of Dolichospermum could be inhibited.
4.2 The effects of temperature increasing on Dolichospermum aggregationTemperature is an important factor affecting the growth of algae (Maheaux et al., 2016), and aromatic amino acid was thought to resist temperature stress, especially at higher temperature (Teng et al., 2019). While high temperatures (27–30 ℃) did harm the formation of large colonies and degraded algae cell proteins, DNA, lipids, and membranes (Zhu et al., 2016; Babele et al., 2017), protein secretion in Dolichospermum was inhibited, which is consistent with the findings of this study. Moreover, the growth of cyanobacteria was related to tryptophan and humic substances in S-EPS, while only tryptophan substances in LB-EPS and TB-EPS (Xu et al., 2013b), the concentration of specific compounds in each type of EPSs were affected by temperature (Wei et al., 2017), which is consistent with the results of this study too.
Temperature influenced the surface characteristics of Dolichospermum, and then affected the aggregation of algae cells. The absolute value of zeta potential can represent the change of surface charge (Liu et al., 2010). The absolute value change of zeta potential showed the same trend as the aggregation rate in this study (Fig. 1c). The contact angles indicated that Dolichospermum showed hydrophilicity under the cultivation temperature. We speculated that it was because Dolichospermum secreted more polysaccharides than protein with the increase in temperature (Fig. 2). The amount of surface charge also altered the aggregation of algae cells, which also confirmed the results of Tan et al. (2018).
In addition, hydrogen bonding, hydrophobic and electrostatic interactions between microorganisms can affect microbial aggregation (Yang et al., 2022). The change of interaction free energy indicated that algae cells tended to bond with water molecules rather than with each other and gradually showed hydrophilicity at high temperature and the Lewis acid-base played a major role in the aggregation of algae cells in process of increasing temperature. The same conclusion was obtained in the study of sludge system (Hou et al., 2015). The results of cell surface thermodynamic analysis further confirmed that the aggregation ability of Dolichospermum changed with temperature.
Dolichospermum acclimated to a higher temperature in natural conditions, usually around 27–30 ℃ (Kłodawska et al., 2019). Dolichospermum increased its metabolic activity and biomass when water temperature increased (Qian et al., 2019). Furthermore, previous research has shown that microcystins are more likely to accumulate and form blooms in the summer (Martinez et al., 2008; Liu et al., 2016), but Dolichospermum blooms have occurred frequently in summer recent years (Olofsson et al., 2019; Qian et al., 2019). This was very different from the aggregation temperature determined in the laboratory. The primary reason could be the single strain culture in the laboratory, as well as the relatively simple environment. As a result, detecting Dolichospermum aggregation is essential, and further researches are needed to focus on how to deal with the aggregation of Dolichospermum and the relationship with the water environment, and how to manage it.
5 CONCLUSIONThe recruitment temperature in this study was 11 ℃ for Dolichospermum growth, and 21 ℃ was the optimum temperature for Dolichospermum. 35 ℃ was the upper limit of temperature tolerance for Dolichospermum growth. The aggregation rate decreased significantly when the cultivation temperature increased from 11 to 37 ℃. The composition and concentration of EPSs affected the cell surface properties of Dolichospermum with temperature change, and thus affected the aggregation of Dolichospermum. During the temperature increase, proteins in EPSs matrix played a major role in promoting the dispersion of algae cells and reducing the aggregation. Surface thermodynamic analysis found that the interfacial free energy and electrostatic repulsion on the surface of algae cells were enhanced, which further increased the dispersion of Dolichospermum.
6 DATA AVAILABILITY STATEMENTAll data generated and/or analyzed during this study are available from the corresponding author upon reasonable request.
7 ACKNOWLEDGMENTWe thank Xiaofeng ZHANG, Jinliu YE, and Caiyu ZOU from Nanjing Normal University for their assistance in samples measurements and field work.
Electronic supplementary material
Supplementary material (Supplementary Table S1 and Figs.S1–S2) is available in the online version of this article at https://doi.org/10.1007/s00343-023-2249-8.
Arévalo S, Nenninger A, Nieves-Morión M, et al. 2021. Coexistence of communicating and noncommunicating cells in the filamentous cyanobacterium Anabaena. mSphere, 6(1): e01091-20.
DOI:10.1128/msphere.01091-20 |
Babele P K, Singh G, Singh A, et al. 2017. UV-B radiation and temperature stress-induced alterations in metabolic events and defense mechanisms in a bloom-forming cyanobacterium Microcystis aeruginosa. Acta Physiologiae Plantarum, 39(1): 248.
DOI:10.1007/s11738-017-2540-4 |
Bahram M, Bro R, Stedmon C, et al. 2006. Handling of Rayleigh and Raman scatter for PARAFAC modeling of fluorescence data using interpolation. Journal of Chemometrics, 20(3-4): 99-105.
DOI:10.1002/cem.978 |
Baker A, Inverarity R. 2004. Protein-like fluorescence intensity as a possible tool for determining river water quality. Hydrological Processes, 18(15): 2927-2945.
DOI:10.1002/hyp.5597 |
Bistgani Z E, Siadat S A, Bakhshandeh A, et al. 2017. Interactive effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenensis celak. The Crop Journal, 5(5): 407-415.
DOI:10.1016/j.cj.2017.04.003 |
Carey C C, Ibelings B W, Hoffmann E P, et al. 2012. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Research, 46(5): 1394-1407.
DOI:10.1016/j.watres.2011.12.016 |
Chen H Z, Zuo S P, Qin B Q, et al. 2019. Research progress in mechanism of Microcystis aggregation and migration. Environmental Science & Technology, 42(1): 142-149.
(in Chinese with English abstract) DOI:10.19672/j.cnki.1003-6504.2019.01.021 |
Dang C Y, Yang Z X, Liu W, et al. 2018. Role of extracellular polymeric substances in biosorption of Pb2+ by a high metal ion tolerant fungal strain Aspergillus niger PTN31. Journal of Environmental Chemical Engineering, 6(2): 2733-2742.
DOI:10.1016/j.jece.2018.04.005 |
Dubois M, Gilles K A, Hamilton J K, et al. 1956. Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28(3): 350-356.
DOI:10.1021/ac60111a017 |
Florence A. 2007. Interfacial forces in aqueous media, C. van Oss, 2nd ed., CRC Press (2006), ISBN: 1-57444-482. International Journal of Pharmaceutics, 331(1): 145.
DOI:10.1016/j.ijpharm.2006.11.036 |
Forni C, Telo' F R, Caiola M G. 1997. Comparative analysis of the polysaccharides produced by different species of Microcystis (Chroococcales, Cyanophyta). Phycologia, 36(3): 181-185.
DOI:10.2216/i0031-8884-36-3-181.1 |
Funke W. 1995. Interfacial forces in aqueous media: by C. J. van Oss, published by Marcel Dekker, New York, 1994, US $ 165. Progress in Organic Coatings, 26(1): 75-76.
DOI:10.1016/0300-9440(95)90006-3 |
Garg R, Maldener I. 2021. The formation of spore-like akinetes: a survival strategy of filamentous cyanobacteria. Microbial Physiology, 31(3): 296-305.
DOI:10.1159/000517443 |
Hadjoudja S, Deluchat V, Baudu M. 2010. Cell surface characterisation of Microcystis aeruginosa and Chlorella vulgaris. Journal of Colloid and Interface Science, 342(2): 293-299.
DOI:10.1016/j.jcis.2009.10.078 |
He J J, Yang P, Zhang W J, et al. 2017. Characterization of changes in floc morphology, extracellular polymeric substances and heavy metals speciation of anaerobically digested biosolid under treatment with a novel chelated-Fe2+ catalyzed Fenton process. Bioresource Technology, 243: 641-651.
DOI:10.1016/j.biortech.2017.06.180 |
Henderson R K, Baker A, Parsons S A, et al. 2008. Characterisation of algogenic organic matter extracted from cyanobacteria, green algae and diatoms. Water Research, 42(13): 3435-3445.
DOI:10.1016/j.watres.2007.10.032 |
Holsinger E C T. 1955. The distribution and periodicity of the phytoplankton of three Ceylon lakes. Journal of Aquatic Ecosystem Stress and Recovery (Formerly Journal of Aquatic Ecosystem Health), 7(1-2): 25-35.
DOI:10.1007/BF00189792 |
Hou X L, Liu S T, Zhang Z T. 2015. Role of extracellular polymeric substance in determining the high aggregation ability of anammox sludge. Water Research, 75: 51-62.
DOI:10.1016/j.watres.2015.02.031 |
Ji C C, Zhou H, Deng S K, et al. 2021. Insight into the adhesion propensities of extracellular polymeric substances (EPS) on the abiotic surface using XDLVO theory. Journal of Environmental Chemical Engineering, 9(6): 106563.
DOI:10.1016/j.jece.2021.106563 |
Kłodawska K, Bujas A, Turos-Cabal M, et al. 2019. Effect of growth temperature on biosynthesis and accumulation of carotenoids in cyanobacterium Anabaena sp. PCC 7120 under diazotrophic conditions. Microbiological Research, 226: 34-40.
DOI:10.1016/j.micres.2019.05.003 |
Kovács A W, Présing M, Vörös L. 2016. Thermal-dependent growth characteristics for Cylindrospermopsis raciborskii (Cyanoprokaryota) at different light availabilities: methodological considerations. Aquatic Ecology, 50(4): 623-638.
DOI:10.1007/s10452-016-9582-3 |
Li X Y, Yang S F. 2007. Influence of loosely bound extracellular polymeric substances (EPS) on the flocculation, sedimentation and dewaterability of activated sludge. Water Research, 41(5): 1022-1030.
DOI:10.1016/j.watres.2006.06.037 |
Liao B Q, Allen D G, Droppo I G, et al. 2001. Surface properties of sludge and their role in bioflocculation and settleability. Water Research, 35(2): 339-350.
DOI:10.1016/S0043-1354(00)00277-3 |
Liu H, Fang H H P. 2002. Characterization of electrostatic binding sites of extracellular polymers by linear programming analysis of titration data. Biotechnology and Bioengineering, 80(7): 806-811.
DOI:10.1002/bit.10432 |
Liu L Z, Huang Q, Qin B Q, et al. 2016. Characterizing cell surface of blooming Microcystis in Lake Taihu, China. Water Science and Technology, 73(11): 2731-2738.
DOI:10.2166/wst.2016.069 |
Liu L Z, Huang Q, Zhang Y L, et al. 2017. Excitation-emission matrix fluorescence and parallel factor analyses of the effects of N and P nutrients on the extracellular polymeric substances of Microcystis aeruginosa. Limnologica, 63: 18-26.
DOI:10.1016/j.limno.2016.10.006 |
Liu L Z, Qin B Q, Zhang Y L, et al. 2014. Extraction and characterization of bound extracellular polymeric substances from cultured pure cyanobacterium (Microcystis wesenbergii). Journal of Environmental Sciences, 26(8): 1725-1732.
DOI:10.1016/j.jes.2014.06.013 |
Liu X M, Sheng G P, Luo H W, et al. 2010. Contribution of extracellular polymeric substances (EPS) to the sludge aggregation. Environmental Science & Technology, 44(11): 4355-4360.
DOI:10.1021/es9016766 |
Liu X M, Sheng G P, Yu H Q. 2007. DLVO approach to the flocculability of a photosynthetic H2-producing bacterium, Rhodopseudomonas acidophila. Environmental Science & Technology, 41(13): 4620-4625.
DOI:10.1021/es070107n |
Maheaux H, Leavitt P R, Jackson L J. 2016. Asynchronous onset of eutrophication among shallow prairie lakes of the northern Great Plains, Alberta, Canada. Global Change Biology, 22(1): 271-283.
DOI:10.1111/gcb.13076 |
Martinez R E, Pokrovsky O S, Schott J, et al. 2008. Surface charge and zeta-potential of metabolically active and dead cyanobacteria. Journal of Colloid and Interface Science, 323(2): 317-325.
DOI:10.1016/j.jcis.2008.04.041 |
Ndlela L L, Oberholster P J, Van Wyk J H, et al. 2016. An overview of cyanobacterial bloom occurrences and research in Africa over the last decade. Harmful Algae, 60: 11-26.
DOI:10.1016/j.hal.2016.10.001 |
Olofsson M, Torstensson A, Karlberg M, et al. 2019. Limited response of a spring bloom community inoculated with filamentous cyanobacteria to elevated temperature and pCO2. Botanica Marina, 62(1): 3-16.
DOI:10.1515/bot-2018-0005 |
Olson J B, Steppe T F, Litaker R W, et al. 1998. N2-fixing microbial consortia associated with the ice cover of Lake Bonney, Antarctica. Microbial Ecology, 36(3): 231-238.
|
Paerl H W, Hall N S, Calandrino E S. 2011. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Science of the Total Environment, 409(10): 1739-1745.
DOI:10.1016/j.scitotenv.2011.02.001 |
Paerl H W, Huisman J. 2009. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environmental Microbiology Reports, 1(1): 27-37.
DOI:10.1111/j.1758-2229.2008.00004.x |
Paerl H W, Paul V J. 2012. Climate change: links to global expansion of harmful cyanobacteria. Water Research, 46(5): 1349-1363.
DOI:10.1016/j.watres.2011.08.002 |
Paerl H W, Priscu J C, Ecol M. 1998. Microbial phototrophic, heterotrophic, and diazotrophic activities associated with aggregates in the permanent ice cover of Lake Bonney, Antarctica. Microbial Ecology, 36(3): 221-230.
DOI:10.1007/s002489900109 |
Qian K M, Dokulil M, Chen Y W. 2019. Do the regular annual extreme water level changes affect the seasonal appearance of Anabaena in Poyang Lake?. PeerJ, 7: e6608.
DOI:10.7717/peerj.6608 |
Qu F S, Liang H, He J G, et al. 2012a. Characterization of dissolved extracellular organic matter (dEOM) and bound extracellular organic matter (bEOM) of Microcystis aeruginosa and their impacts on UF membrane fouling. Water Research, 46(9): 2881-2890.
DOI:10.1016/j.watres.2012.02.045 |
Qu F S, Liang H, Wang Z Z, et al. 2012b. Ultrafiltration membrane fouling by extracellular organic matters (EOM) of Microcystis aeruginosa in stationary phase: influences of interfacial characteristics of foulants and fouling mechanisms. Water Research, 46(5): 1490-1500.
DOI:10.1016/j.watres.2011.11.051 |
Reddy Y P, Yadav R K, Tripathi K, et al. 2019. Isolation and characterization of high temperature tolerant mutant from the cyanobacterium Anabaena doliolum. Journal of Basic Microbiology, 59(3): 314-322.
DOI:10.1002/jobm.201800447 |
Richert L, Golubic S, Le Guédès R, et al. 2005. Characterization of exopolysaccharides produced by cyanobacteria isolated from Polynesian microbial mats. Current Microbiology, 51(6): 379-384.
DOI:10.1007/s00284-005-0069-z |
Sakharova E G, Krylov A V, Sabitova R Z, et al. 2020. Horizontal and vertical distribution of phytoplankton in the alpine Lake Sevan (Armenia) during the summer cyanoprokaryota bloom. Contemporary Problems of Ecology, 13: 60-70.
DOI:10.1134/S1995425520010072 |
Sheng G P, Yu H Q, Li X Y. 2010. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: a review. Biotechnology Advances, 28(6): 882-894.
DOI:10.1016/j.biotechadv.2010.08.001 |
Srivastava A, Biswas S, Yadav S, et al. 2021. Acute cadmium toxicity and post-stress recovery: insights into coordinated and integrated response/recovery strategies of Anabaena sp. PCC 7120. Journal of Hazardous Materials, 411: 124822.
DOI:10.1016/j.jhazmat.2020.124822 |
Sukenik A, Rücker J, Maldener I. 2019. Dormant cells (akinetes) of filamentous cyanobacteria demonstrate a great variability in morphology, physiology, and ecological function. In: Mishra A K, Tiwari D N, Rai A N eds. Cyanobacteria: From Basic Science to Applications. Elsevier, Amsterdam. p. 65-77, https://doi.org/10.1016/B978-0-12-814667-5.00004-0.
|
Swe T, Miles C O, Cerasino L, et al. 2021. Microcystis, Raphidiopsis raciborskii and Dolichospermum smithii, toxin producing and non-toxigenic cyanobacteria in Yezin Dam, Myanmar. Limnologica, 90: 125901.
DOI:10.1016/j.limno.2021.125901 |
Tan L R, Xia P F, Zeng R J, et al. 2018. Low-level concentrations of aminoglycoside antibiotics induce the aggregation of cyanobacteria. Environmental Science and Pollution Research, 25(17): 17128-17136.
DOI:10.1007/s11356-018-1894-5 |
Tan X, Shu X Q, Duan Z P, et al. 2020. Two types of bound extracellular polysaccharides and their roles in shaping the size and tightness of Microcystis colonies. Journal of Applied Phycology, 32: 255-262.
DOI:10.1007/s10811-019-01937-z |
Teng Z D, Shao W, Zhang K Y, et al. 2019. Pb biosorption by Leclercia adecarboxylata: protective and immobilized mechanisms of extracellular polymeric substances. Chemical Engineering Journal, 375: 122113.
DOI:10.1016/j.cej.2019.122113 |
Vogelaar J C T, De Keizer A, Spijker S, et al. 2005. Bioflocculation of mesophilic and thermophilic activated sludge. Water Research, 39(1): 37-46.
DOI:10.1016/j.watres.2004.07.027 |
Wang L F, Li Y, Wang L, et al. 2018. Responses of biofilm microorganisms from moving bed biofilm reactor to antibiotics exposure: protective role of extracellular polymeric substances. Bioresource Technology, 254: 268-277.
DOI:10.1016/j.biortech.2018.01.063 |
Wei D, Yan T, Zhang K Y, et al. 2017. Qualitative and quantitative analysis of extracellular polymeric substances in partial nitrification and full nitrification reactors. Bioresource Technology, 240: 171-176.
DOI:10.1016/j.biortech.2017.02.115 |
Weiss G L, Kieninger A K, Maldener I, et al. 2019. Structure and function of a bacterial gap junction analog. Cell, 178(2): 374-384.e15.
DOI:10.1016/j.cell.2019.05.055 |
Wen X G, Gong H M, Lu C M. 2005. Heat stress induces an inhibition of excitation energy transfer from phycobilisomes to photosystem Ⅱ but not to photosystem I in a cyanobacterium Spirulina platensis. Plant Physiology and Biochemistry, 43(4): 389-395.
DOI:10.1016/j.plaphy.2005.03.001 |
Xing L L, Yang J X, Ni B J, et al. 2022. Insight into the generation and consumption mechanism of tightly bound and loosely bound extracellular polymeric substances by mathematical modeling. Science of the Total Environment, 811: 152359.
DOI:10.1016/j.scitotenv.2021.152359 |
Xu H C, Cai H Y, Yu G H, et al. 2013a. Insights into extracellular polymeric substances of cyanobacterium Microcystis aeruginosa using fractionation procedure and parallel factor analysis. Water Research, 47(6): 2005-2014.
DOI:10.1016/j.watres.2013.01.019 |
Xu H C, He P J, Wang G Z, et al. 2010. Three-dimensional excitation emission matrix fluorescence spectroscopy and gel-permeating chromatography to characterize extracellular polymeric substances in aerobic granulation. Water Science and Technology, 61(11): 2931-2942.
DOI:10.2166/wst.2010.197 |
Xu H C, Jiang H L, Yu G H, et al. 2014. Towards understanding the role of extracellular polymeric substances in cyanobacterial Microcystis aggregation and mucilaginous bloom formation. Chemosphere, 117: 815-822.
DOI:10.1016/j.chemosphere.2014.10.061 |
Xu H C, Yu G H, Jiang H L. 2013b. Investigation on extracellular polymeric substances from mucilaginous cyanobacterial blooms in eutrophic freshwater lakes. Chemosphere, 93(1): 75-81.
DOI:10.1016/j.chemosphere.2013.04.077 |
Yang F, Li H X, Wang S, et al. 2022. Differences in exopolysaccharides of three microbial aggregates. Environmental Technology, 43(19): 2909-2921.
DOI:10.1080/09593330.2021.1909658 |
Yang Z, Kong F X, Shi X L, et al. 2008. Changes in the morphology and polysaccharide content of Microcystis aeruginosa (cyanobacteria) during flagellate grazing. Journal of Phycology, 44(3): 716-720.
DOI:10.1111/j.1529-8817.2008.00502.x |
Yu G H, He P J, Shao L M. 2009. Characteristics of extracellular polymeric substances (EPS) fractions from excess sludges and their effects on bioflocculability. Bioresource Technology, 100(13): 3193-3198.
DOI:10.1016/j.biortech.2009.02.009 |
Zhang Q, Yu S C, Wang Q, et al. 2021. Quantitative proteomics reveals the protein regulatory network of Anabaena sp. PCC 7120 under nitrogen deficiency. Journal of Proteome Research, 20(8): 3963-3976.
DOI:10.1021/acs.jproteome.1c00302 |
Zhu W, Zhou X H, Chen H M, et al. 2016. High nutrient concentration and temperature alleviated formation of large colonies of Microcystis: evidence from field investigations and laboratory experiments. Water Research, 101: 167-175.
DOI:10.1016/j.watres.2016.05.080 |